首页 > 疾病防控/ 正文
深度解析医学证据,lxfs.net为你支撑决策
摘要
帕金森病(PD)伴神经原性直立性低血压(nOH)的病理机制正由外周交感神经去支配向中枢网络失调演变。本文探讨脑干蓝斑(LC)亚区退化及中枢自主网络(CAN)与躯体-认知动作网络(SCAN)异常在nOH发生中的核心作用。当前研究难点在于界定网络超连通性是生理代偿还是病理标志,且缺乏纵向多模态影像数据。基于CAN、SCAN障碍的全新认知,PD伴nOH的治疗正转向靶向皮质节点、脊髓硬膜外电刺激(eSCS)及闭环神经调控等精准干预范式,这为临床个体化管理提供新视角。
关键词:帕金森病;神经原性直立性低血压;自主神经网络;脑干蓝斑;神经调控
Key words:Parkinson disease;Neurogenic orthostatic hypotension;Autonomic network;Locus coeruleus;Neuromodulation
帕金森病(PD)现已被重新定义为一种累及全身多系统的复杂神经系统变性疾病,而非单纯的中枢神经系统运动障碍[1,2]。在众多的非运动症状(non-motor symptom, NMS)中,自主神经功能障碍尤为普遍且隐匿。其中,神经原性直立性低血压(neurogenic orthostatic hypotension, nOH)作为心血管自主神经衰竭最具代表性的表型,在PD患者群体中的患病率高达30%~50%,极易诱发晕厥、跌倒等严重不良事件,显著削弱患者的预期寿命与生活质量[3,4]。究其病理根源,近年来学术界对PD合并nOH的机制认知正经历一场深刻的范式转移,本文将从PD合并nOH的中枢机制与精准神经调控治疗最新进展方面做系统综述。
1 外周神经系统机制的局限性
长期以来,临床对PD合并nOH的病理生理机制解释,多局限于外周交感神经系统的去支配作用。组织病理学研究确证,α-突触核蛋白(α-synuclein,α-syn)广泛沉积于皮肤、子宫颈及下肢自主神经纤维内[4-7]。尸检与免疫荧光技术亦直观呈现了外周交感去甲肾上腺素能纤维的显著脱失[8,9]。与之相印证,碘-123标记的间碘苄胍心肌显像(iodine-123 metaiodobenzylguanidine myocardial imaging,123I-MIBG)清晰揭示了心脏交感神经的早期去支配改变[10-13]。基于上述广泛的外周病理沉积现象,当前靶向α-syn的单抗药物(如Prasinezumab,PADOVA研究)正试图从源头阻断病理蛋白扩布,以期延缓疾病进展[14]。
然而,单纯的外周交感神经损伤或骨骼肌泵功能减退,难以全盘解释nOH临床表型的高度异质性。近期生理学研究敏锐捕捉到了这一矛盾:低肌肉质量(low muscle mass,LMM)在PD患者中极为普遍(患病率达35.0%),并可诱发站立后30 s内的早期收缩压(systolic blood pressure,SBP)骤降,但LMM与临床确诊的nOH患病率及直立不耐受症状的严重程度间并未展现出统计学相关性(P=0.412),该发现有力提示:仅凭外周血管阻力降低或骨骼肌泵功能受损等周围性因素,尚不足以驱动严重且持续的临床nOH发生[15]。由此推演,在PD复杂的病理演进轨迹中,可能潜藏着更高层级的中枢自主神经调控网络代偿失调[15,16]。
2 脑干蓝斑角色的重新认识
在探寻中枢机制的进程中,作为中枢去甲肾上腺素(norepinephrine,NA)主要源泉的脑干蓝斑(locus coeruleus,LC)一度被确立为核心靶点。LC-NA系统在调节觉醒、应激反应及自主神经稳态中扮演着不可或缺的角色[17-22]。传统Braak病理分级体系亦指出,LC是PD病理累及极早的解剖区域,其神经元脱失程度甚至可能超越黑质致密部[18,23-27]。伴随神经黑色素敏感磁共振成像(neuromelanin-sensitive magnetic resonance imaging,NM-MRI)技术的临床应用,高分辨率影像学证据对传统的“LC中心论”构成了严峻挑战。Palermo等[24]的队列研究揭示,尽管PD患者整体LC信号强度显著劣于健康对照组,但在伴发OH(PDOH+)与不伴发OH(PDOH-)的亚组之间,LC的各项MRI测值间差异无统计学意义。究其原因,Palermo等[24]的研究使用了3.0T MRI测量LC的整体信号,这种广泛性的测量方式极易掩盖并能稀释特定亚区的局部病理特征[21,24]。
与之相反,Madelung等[28]独立研究明确证实,较小的LC体积与直立位收缩压SBP的剧烈下降紧密相关(ρ=0.42,P=0.003)。面对上述影像与临床表型相悖的困境,最新7.0T超高场强MRI研究提供了神经解剖学依据:LC内部实则存在高度特异性的头尾、背腹侧拓扑梯度(rostro-caudal gradient)。具体而言,尾侧LC主要发出下行纤维投射至脊髓,负责调节自主神经与血压,其特异性退化与PD合并nOH的严重程度呈高度正相关,而头侧(rostral)则主要投射至前脑[28]。此外,Beach等[21]最新研究亦指出,LC变性在多系统萎缩(multiple system atrophy,MSA)与PD间的鉴别诊断价值需结合亚区分析方能显现。
进一步剖析其微观机制,目前的“胞体-突触”变性失步理论提供了一个可能的视角:相较于反映细胞体完整性的MRI信号,表征突触末梢完整性的11C-MeNER PET信号与nOH的临床关联更为紧密[24,29]。该现象强烈提示,LC系统的功能衰竭极可能是从远端轴突起始的逆行性病变[24,30]。这一系列探索表明,单一核团的局灶性病变远不足以驱动机制错综复杂的nOH,中枢自主神经调节的全面失代偿,势必牵涉更高层级中枢自主网络(central autonomic network,CAN)的全局性功能减退[16,24](见图1)。
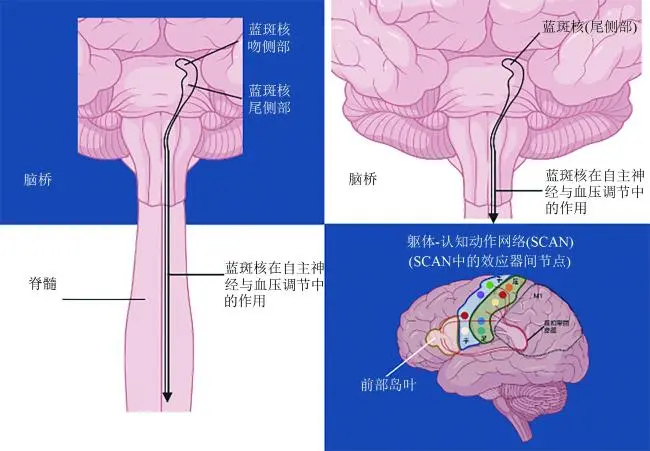
注:LC,蓝斑;SCAN,躯体-认知动作网络。
图1 LC梯度与SCAN枢纽的“整合模型”
3 中枢自主网络与躯体-认知动作网络的异常
顺应上述理论推演,神经影像学研究的焦点正式由“外周、脑干去支配”向“皮质网络调控失灵”维度跃迁。多项证据表明,负责“自上而下”调控的CAN及其与突显网络(salience network,SN)的交互异常,在nOH的病理演进中可能占据了核心枢纽地位[15,16]。
3.1 突显网络与结构协变网络的拓扑解体
在伴发nOH的PD队列中,以岛叶前部和前扣带回为核心的突显网络内部节点连通性呈现显著衰减,且左侧吻侧前额叶皮质(rostral prefrontal cortex,RPFC)与右侧前脑岛之间的连通强度,与SBP的直立位下降幅度呈显著负相关[16,31]。深入至拓扑组织层面,基于灰质体积的结构协变网络(structural covariance network,SCN)分析证实,高自主神经症状负荷患者的大脑小世界指数(σ)及归一化集群系数(γ)均发生断崖式下降。作为CAN核心“桥梁”的右侧丘脑节点中心度(betweenness centrality,BC)显著降低,标志着全脑信息整合能力的实质性破坏[32,33]。与之相对应的代偿现象是,基于图论的网络分析捕捉到左侧中颞回(left middle temporal gyrus)节点效率(nodal efficiency)的异常攀升,这被普遍解读为大脑应对CAN结构损伤的局部网络重塑与代偿反应[32]。
3.2 躯体-认知动作网络的提出与病理生理的重塑
Gordon等[34]利用高精度功能磁共振成像(high-resolution functional magnetic resonance imaging,HRFM)技术对Penfield经典的运动皮质小人(primary motor cortex,M1)模型进行了重构,提出了“集成-隔离”模型:M1并非单一连续体,而是由效应器特异性区域(负责隔离精细运动)与躯体-认知动作网络(somato-cognitive action network,SCAN)交替镶嵌构成[35]。解剖学上,SCAN与扣带岛盖网络(cingulo-opercular network,CON)保持着强烈的全脑功能连通性,在执行动作规划、轴向躯体运动以及自主神经调控(如血压、心率稳态维持)时被同步激活[34-36]。该理论的提出为阐释PD临床表型的高度异质性提供了统一的神经生物学底座。Ren等[35]的最新研究发现,PD的核心受累区域(如黑质、壳核)及传统脑深部电刺激(deep brain stimulation,DBS)靶点,均展现出与SCAN的高度选择性连接。该研究进一步证实,SCAN与皮质下核团的超连通性(hyperconnectivity)与患者的运动控制障碍、认知衰退以及自主神经、唤醒功能障碍(autonomic/arousal dysfunction)呈显著正相关。映射至临床实践,强直-少动型(akinetic-rigid,AR)患者往往伴随更为严重的轴向运动障碍(即SCAN核心功能区受损),nOH患病率亦显著攀升,从侧面提示了该亚型符合“由下而上”(body-first)的病理扩布特征[37,38]。反观震颤为主型(tremor dominant,TD)患者,其病理较少波及SCAN节点,nOH发生率相应处于低位。将PD概念化为一种“SCAN障碍”,为深刻理解运动症状与nOH等非运动症状的病理交织提供了极具价值的理论框架[9,34,35]。
4 超连通性的作用标志
CAN、SCAN网络异常的剖析引发出一项核心学术争议:疾病早期所观测到的网络超连通性,究竟是大脑应对外周去支配损伤的生理性代偿,还是驱动疾病恶化的病理性标志?
4.1 代偿假说
当外周自主神经遭遇去支配打击时,中枢系统会自发上调内部连通性以维系生理稳态。Conti等[39]借助高密度脑电图(high-density electroencephalography,HD-EEG)证实,伴有中重度自主神经症状的早期(early-stage)PD患者,在α频段内表现出岛叶与边缘系统的功能超连通性,此现象被其界定为针对外周损伤的中枢代偿机制。同理,前述结构网络分析中观测到的小脑蚓部(vermis)及左侧中颞回节点效率的异常增加,亦被视作对丘脑“桥梁”塌陷在拓扑层面的中枢代偿[32]。
4.2 病理假说
Ren等[35]发现PD患者皮质下关键核团与皮质SCAN之间存在高度特异性的超连通性,且该连通强度与患者的运动障碍、认知衰退及自主神经功能障碍呈显著正相关。从电生理机制审视,此类超连通性往往伴生β频段(13~35 Hz)的异常同步化,实质上阻滞了神经网络的灵活动态调整[34,40,41]。最有力的反证在于:当施以有效的神经调控干预[如DBS、重复经颅磁刺激(repetitive transcranial magnetic stimulation,rTMS)]或左旋多巴替代治疗后,该超连通性被显著抑制(下调),患者临床症状随之改善。这一因果链条提示,超连通性本身即为疾病的病理状态(过度耦合),打破这种病理锁定才是获取临床获益的关键[35]。
4.3 争议的整合与演进
梳理现有文献脉络,上述分歧可能源于观测频段与疾病演进阶段的异质性。当前证据链倾向于支持一种频段特异性的动态演变模型:α频段的超连通性可能更多介入疾病极早期的功能代偿[39],而β频段或低频[功能磁共振成像(functional magnetic resonance imaging,fMRI)]的超连通性,则表征了伴随病理蛋白广泛扩布而引发的失控与病理性环路锁定[23,35,42]。必须指出的是,由于目前缺乏对同一队列从前驱期[如单纯快速眼动睡眠行为障碍(rapid eye movement sleep behavior disorder,RBD)阶段]到中晚期PD合并nOH的纵向高频脑电、fMRI追踪数据,所以尚无法精准界定网络从“生理代偿”滑向“过度耦合(病理锁定)”的精确时间节点或生物标志物临界值(biomarker cutoff)[35]。
5 走向精准神经调控治疗
基于“PD及PD合并nOH核心为SCAN、CAN网络障碍”的全新病理认知,临床治疗策略正跨越单纯的外周容量扩充与血管活性药物补充,迈向针对中枢或脊髓环路的精准神经调控干预。
5.1 精准皮质调控:靶向SCAN节点的无创干预
最新功能影像学证据提示,直接靶向SCAN皮质节点实施干预,较之传统初级运动区刺激可获得更为卓越的临床获益。特别是经颅磁刺激(transcranial magnetic stimulation,TMS)与低强度聚焦超声(focused ultrasound,FUS)等无创神经调控技术的发展,使得靶向SCAN网络的干预窗口得以大幅前移[35,43]。一项随机对照试验表明,依托个性化功能连接图谱精准定位SCAN节点并施以rTMS、间歇性脉θ冲刺激(intermittent θ burst stimulation, iTBS),其改善PD整体运动症状的效能,较传统效应器区域刺激显著提升[35]。与此同时,皮质脑电图(electrocorticography,ECoG)记录亦证实,DBS的最优刺激靶点在功能拓扑上,恰好与SCAN皮质节点呈现紧密耦合[33]。
5.2 脊髓硬膜外电刺激:重塑难治性nOH的血流动力学
面对常规药物难治的重度nOH,脊髓硬膜外电刺激(epidural electrical spinal cord stimulation,eSCS)技术展现出了令患者从“卧床不起”跨越至“自主行走”的巨大转化潜能。Zhu等[44]的临床报道显示,在接受T10-T11节段eSCS治疗后,重度nOH患者的SBP直立位骤降幅度获得显著纠正,其独立行走能力及6分钟步行试验(6-min walk test,6MWT)指标均实现大幅改善。探究其解剖学机制,有效电刺激场必须精准覆盖位于下胸段(T11-T13)的“血流动力学热点(hemodynamic hotspots)”。该概念首创于洛桑联邦理工学院(École Polytechnique Fédérale de Lausanne,EPFL)团队,其核心机制在于逆向激活内脏交感神经节的后根传入纤维,从而为eSCS的强效升压效应奠定了解剖学基石[45]。反之,若刺激靶点偏离至腰骶部,不仅升压效应微弱,更可能诱发自主神经反射异常[46,47]。
5.3 闭环控制与 “智能自主神经义肢”的展望
为破解开放环路固定刺激易引发的过度代偿或疗效衰减难题,闭环神经调控(closed-loop neuromodulation)已跃升为该领域的核心演进方向。针对此前闭环系统易受运动伪影和心电信号污染的瓶颈,Gandhi等[47]成功开发出基于比例-积分-微分(proportional-integral-derivative,PID)控制器的患者优化闭环eSCS系统。该系统通过连续血压监测进行实时动态调整,不仅有效改善了收缩压的剧烈波动,还通过算法优化大幅降低了植入设备的功耗,初步证明了闭环策略在真实世界复杂电磁环境下克服血流动力学波动的可行性[47,48]。
尽管前景广阔,闭环神经调控在当前临床转化中仍面临严峻的技术壁垒。通过DBS对其皮质节点进行逆向调控展现出巨大潜力。然而,目前的探索多集中于利用高精度MRI绘制皮质下结构与SCAN网络的异常连接图谱。学界仍需开展更多前瞻性临床试验,为将来SCAN作为精准调控靶点提供直接的安全性与有效性循证依据[35,49]。在这些大样本队列研究完成前,仍需关注其潜在的脱靶效应(off-target effects)。
着眼于临床综合管理,伴随神经调控与靶向药物的迭代,2024年欧洲心脏病学会(European Society of Cardiology,ESC)最新指南亦明确强调,针对合并nOH的高血压群体,应摒弃僵化的降压标准,采取更为灵活的个体化血压管理目标,以期在改善直立耐受性与遏制心血管终点事件风险之间,取得最佳的临床平衡[50]。
6 结语
PD伴nOH的机制认知正经历从“外周去支配”向“中枢网络失调”的深刻范式转移。LC亚区特异性退化与CAN、SCAN网络的拓扑异常,共同构成了nOH发生的中枢病理基础。然而,网络超连通性究竟是代偿还是致病标志,仍有赖于纵向多模态影像与高频电生理研究的进一步验证。基于上述研究,当前治疗策略已从单纯的血管活性药物补充,拓展至靶向SCAN节点的无创皮质调控、eSCS及闭环神经调控等精准干预模式。未来研究应聚焦于3个方向:(1)界定网络超连通性从代偿滑向病理的临界点与生物标志物;(2)开展大样本前瞻性临床试验,验证SCAN作为精准调控靶点的有效性与安全性;(3)推动闭环自适应系统的临床转化,最终实现基于中枢自主神经网络的个体化综合管理。
参考文献
[1] Mazzotta GM, Conte C. Alpha synuclein toxicity and non-motor Parkinson’s[J]. Cells, 2024, 13(15): 1265.
[2] 崔诗爽, 张冬燕, 陈先文, 等. 帕金森病血压管理专家共识(第二版)[J]. 中华高血压杂志, 2023, 31(9): 809-820.
[3] Velseboer DC, de Haan RJ, Wieling W, et al. Prevalence of orthostatic hypotension in Parkinson’s disease: A systematic review and meta-analysis[J]. Park Relat Disord, 2011, 17(10): 724-729.
[4] Palma JA, Kaufmann H. Orthostatic hypotension in Parkinson disease[J]. Clin Geriatr Med, 2020, 36(1): 53-67.
[5] Sharabi Y, Vatine GD, Ashkenazi A. Parkinson’s disease outside the brain: Targeting the autonomic nervous system[J]. Lancet Neurol, 2021, 20(10): 868-876.
[6] Gibbons CH, Levine T, Adler C, et al. Skin biopsy detection of phosphorylated α-synuclein in patients with synucleinopathies[J]. JAMA, 2024, 331(15): 1298-1306.
[7] Coughlin DG, Adler CH, Barbosa W, et al. CSF α-synuclein seed amplification assays and skin immunofluorescence: Clinical applications, research opportunities, and knowledge gaps[J]. Neurology, 2026, 106(3): e214648.
[8] Isonaka R, Sullivan P, Goldstein DS. Pathophysiological significance of increased α-synuclein deposition in sympathetic nerves in Parkinson’s disease: A post-mortem observational study[J]. Transl Neurodegener, 2022, 11(1): 15.
[9] Grosu C, Noea O, Mastaleru A, et al. Neurogenic orthostatic hypotension in Parkinson disease: A narrative review of diagnosis and management[J]. J Clin Med, 2025, 14(2): 630.
[10] Amino T, Orimo S, Itoh Y, et al. Profound cardiac sympathetic denervation occurs in Parkinson disease[J]. Brain Pathol, 2006, 15(1): 29-34.
[11] Orimo S, Uchihara T, Nakamura A, et al. Axonal alpha-synuclein aggregates herald centripetal degeneration of cardiac sympathetic nerve in Parkinson’s disease[J]. Brain, 2008, 131(Pt 3): 642-650.
[12] Clement G, Cavillon G, Vuillier F, et al. Unveiling autonomic failure in synucleinopathies: Significance in diagnosis and treatment[J]. Rev Neurol, 2024, 180(1-2): 79-93.
[13] Ryu DW, Yoo SW, Oh Y, et al. Cardiac 123I-meta-iodobenzylguanidine imaging as a biomarker for body-first Parkinson’s disease: Linking peripheral α-synuclein to clinical subtyping[J]. J Mov Disord, 2026, 19(1): 1-10.
[14] Nikolcheva T, Pagano G, Pross N, et al. A Phase 2b, multicenter, randomized, double-blind, placebo-controlled study to evaluate the efficacy and safety of intravenous prasinezumab in early-stage Parkinson’s disease (PADOVA): Rationale, design, and baseline data[J]. Park Relat Disord, 2025, 132: 107257.
[15] Choi S, Kim R, Kwon S, et al. Prevalence of low muscle mass and its association with orthostatic hypotension and related symptoms in Parkinson’s disease[J]. npj Park Dis, 2026, 12: 41.
[16] de Lima Xavier L, Dayan E, Browner N, et al. Evidence of central autonomic dysfunction in orthostatic hypotension of Parkinson’s disease[J]. Park Relat Disord, 2025, 141: 108093.
[17] Poe GR, Foote S, Eschenko O, et al. Locus coeruleus: A new look at the blue spot[J]. Nat Rev Neurosci, 2020, 21(11): 644-659.
[18] Paredes-Rodriguez E, Vegas-Suarez S, Morera-Herreras T, et al. The noradrenergic system in Parkinson’s disease[J]. Front Pharmacol, 2020, 11: 435.
[19] Grimm C, Duss SN, Privitera M, et al. Tonic and burst-like locus coeruleus stimulation distinctly shift network activity across the cortical hierarchy[J]. Nat Neurosci, 2024, 27(11): 2167-2177.
[20] Caestecker S, Lescrauwaet E, Boon P, et al. The locus coeruleus: Noradrenergic system in the healthy and diseased brain: A narrative review[J]. Euro J Neurology, 2025, 32(9): e70337.
[21] Beach PA, Hyland S, Chen X, et al. Locus coeruleus degeneration is associated with orthostatic hypotension in Parkinson’s disease and multiple system atrophy[J]. Clin Auton Res, 2026, 36(1): 53-64.
[22] Sayegh ALC, Janzen A, Strzedulla I, et al. Increased muscle sympathetic nerve activity and impaired baroreflex control in isolated REM-sleep behavior disorder[J]. Clin Neurophysiol, 2021, 132(7): 1537-1542.
[23] Braak H, Del Tredici K, Rüb U, et al. Staging of brain pathology related to sporadic Parkinson’s disease[J]. Neurobiol Aging, 2003, 24(2): 197-211.
[24] Palermo G, Galgani A, Bellini G, et al. Neurogenic orthostatic hypotension in Parkinson’s disease: Is there a role for locus coeruleus magnetic resonance imaging?[J]. J Neural Transm, 2024, 131(2): 157-164.
[25] Miki Y, Foti SC, Hansen D, et al. Hippocampal α-synuclein pathology correlates with memory impairment in multiple system atrophy[J]. Brain, 2020, 143(6): 1798-1810.
[26] Isaias IU, Trujillo P, Summers P, et al.Neuromelanin imaging and dopaminergic loss in Parkinson’s disease[J]. Front Aging Neurosci, 2016, 8: 196.
[27] Zarow C, Lyness SA, Mortimer JA, et al. Neuronal loss is greater in the locus coeruleus than nucleus basalis and substantia nigra in Alzheimer and Parkinson diseases[J]. Arch Neurol, 2003, 60(3): 337.
[28] Madelung CF, Meder D, Fuglsang SA, et al. Locus coeruleus shows a spatial pattern of structural disintegration in Parkinson’s disease[J].Mov Disord, 2022, 37(3): 479-489.
[29] Sommerauer M,Fedorova TD, Hansen AK, et al. Evaluation of the noradrenergic system in Parkinson’s disease: An 11C-MeNER PET and neuromelanin MRI study[J]. Brain,2018,141(2): 496-504.
[30] Cheng HC, Ulane CM, Burke RE. Clinical progression in Parkinson disease and the neurobiology of axons[J]. Ann Neurol, 2010, 67(6): 715-725.
[31] Menon V, Uddin LQ, Saliency, switching, attention and control: A network model of insula function[J]. Brain Struct Funct, 2010. 214(5-6): 655-67.
[32] Zhang H, Huang Y, Lu Q, et al. Abnormal topological organization of structural covariance networks in early-stage Parkinson’s disease patients with autonomic dysfunction[J]. Quant Imaging Med Surg, 2025, 15(11): 10502-10515.
[33] Bassett DS, Bullmore ET.Small-world brain networks revisited[J]. Neuroscientist, 2017,23(5): 499-516.
[34] Gordon EM, Chauvin RJ,Van AN, et al.A somato-cognitive action network alternates with effector regions in motor cortex[J]. Nature, 2023, 617(7960): 351-359.
[35] Ren J, Zhang W, Dahmani L, et al. Parkinson’s disease as a somato-cognitive action network disorder[J]. Nature, 2026, 651(8107): 1030-1038.
[36] Dosenbach NUF, Visscher KM, Palmer ED, et al. A core system for the implementation of task sets[J]. Neuron, 2006, 50(5): 799-812.
[37] Oikonomou P, Koschel J, Altmann CF, et al. Differences in prevalence of orthostatic hypotension between Parkinson’s Disease motor subtypes[J]. Neurol Neurochir Pol, 2025, 59(2): 188-190.
[38] Horsager J, Borghammer P. Brain-first vs. body-first Parkinson’s disease: An update on recent evidence[J]. Park Relat Disord, 2024, 122: 106101.
[39] Conti M, Garasto E, Bovenzi R, et al. Insular and limbic abnormal functional connectivity in early-stage Parkinson’s disease patients with autonomic dysfunction[J]. Cereb Cortex, 2024, 34(7): bhae270.
[40] 李秋儒,李杭,王迪,等.帕金森病直立位低血压与认知障碍的关系[J].中风与神经疾病杂志,2025,42(9):861-864.
[41] Cagnan H, Pedrosa D, Little S, et al. Stimulating at the right time: Phase-specific deep brain stimulation[J]. Brain, 2017, 140(1): 132-145.
[42] Mantini D, Perrucci MG, Del Gratta C, et al. Electrophysiological signatures of resting state networks in the human brain[J]. Proc Natl Acad Sci U S A, 2007, 104(32): 13170-13175.
[43] Vuong MQ, Sreenivasan V, Lee S, et al. Low-intensity transcranial ultrasound effects on the ventral intermediate nucleus and zona incerta in Parkinson’s disease tremor[J]. Brain Stimul, 2026, 19(1): 103025.
[44] Zhu X, Tan Y, Gu Z, et al. Epidural spinal cord stimulation for the management of orthostatic hypotension in Parkinson’s disease: A case report[J]. Brain Stimul, 2025, 18(5): 1370-1372.
[45] Squair JW, Berney M, Castro Jimenez M, et al. Implanted system for orthostatic hypotension in multiple-system atrophy[J]. N Engl J Med, 2022, 386(14): 1339-1344.
[46] Phillips AA, Gandhi AP, Hankov N, et al. An implantable system to restore hemodynamic stability after spinal cord injury[J]. Nat Med, 2025, 31(9): 2946-2957.
[47] Gandhi AP, Hankov N, Intering N, et al. PID controlled epidural electrical stimulation for managing orthostatic hypotension in individuals with spinal cord injury[C]//47th Annual International Conference of the IEEE Engineering in Medicine and Biology Society (EMBC 2025). Copenhagen, Denmark: IEEE Engineering in Medicine and Biology Society (EMBS), 2025: 1-7.
[48] Cho M, Choo M, Koh M, et al. Neuroprosthetic closed-loop strategy for sustained blood pressure reduction via simultaneous stimulation and recording from the spinal cord[J]. Neurotherapeutics, 2025, 22(6): e00758.
[49] Neumann WJ, Gilron R, Little S, et al.Adaptive deep brain stimulation: From experimental evidence toward practical implementation[J]. Mov Disord, 2023, 38(6): 937-948.
[50] McEvoy JW, McCarthy CP, Bruno RM, et al.2024 ESC Guidelines for the management of elevated blood pressure and hypertension[J]. Eur Heart J, 2024. 45(38): 3912-4018.
引证本文
谢紫馨 , 马梓滨 , 赵静 , 王馨悦 , 马少辰 , 石翀 , 王培福 , 李继来 , 杨静 , 杜继臣 , 万志荣. 帕金森病自主神经障碍的中枢机制与精准神经调控治疗研究进展[J]. 中风与神经疾病杂志, 2026, 43(5): 407-411.
小提示:本篇资讯仅在梅斯医学APP中开放阅读,请扫描二维码直接下载APP
