首页 > 医疗新闻/ 正文
深度解析医学证据,lxfs.net为你支撑决策
【论著】
本研究通过生物信息学与动物实验分析,对机械通气相关性肺损伤(VILI)中自噬通量的变化及其机制进行深入探讨,旨在证实VILI中存在自噬通量障碍的现象并筛选潜在靶点,为干预VILI提供新的思路和策略。
1 材料与方法
1.1 生物信息学分析
1.1.1 数据集来源
从基因表达综合数据库(GEO)中下载数据集GSE11434,从小鼠基因组信息数据库和人类自噬数据库(HADb)中获得695个自噬相关基因(ARGs)。GSE11434芯片数据集基于GPL1261平台,该数据集的样本来源于10只小鼠:其中5只为大潮气量通气组,以20 ml/kg呼吸机通气3 h构建VILI模型;另外5只为非通气组,自主呼吸3 h。干预结束后,原研究者取上述10只小鼠的肺组织,基于Affymetrix 430 2.0 阵列平台进行了全基因组信使RNA(mRNA)表达谱测序,本研究从中获取了该测序数据用于后续分析。
1.1.2 差异表达基因(DEGs)的筛选
使用R软件(4.2.1)版本的Limma包,筛选VILI组(对应原数据集大潮气量通气组)与对照组(Con组,对应原数据集非通气组)小鼠肺组织中的DEGs。在P<0.05和|log2FC|≥0.58的条件下筛选DEGs,log2FC>0的DEGs被认为上调,而log2FC<0的DEGs被认为下调。用R软件的ggplot2[3.4.4]包绘制主成分分析(PCA)图,以评估组内样本的生物学重复性及组间的差异性;用ggplot2[3.4.4]包绘制箱线图,通过观察各样本基因表达量的分布中位线,评估样本间的一致性;用ggplot2[3.4.4]包绘制火山图,以直观展示DEGs的整体分布及差异情况;利用ComplexHeatmap[2.13.1]包绘制聚类热图,用于评估组内样本的表达一致性及组间的差异模式。
1.1.3 自噬相关差异表达基因(DEARGs)的识别
分别将上调的DEGs和下调的DEGs与695个ARGs进行交集分析,并利用ggplot2[3.4.4]和VennDiagram[1.7.3]包绘制韦恩图,以识别VILI相关的DEARGs。利用ComplexHeatmap[2.13.1]包绘制聚类热图,分析DEARGs的相对表达模式。
1.1.4 DEARGs功能富集分析
利用R软件的clusterProfiler[4.4.4]包对DEARGs进行功能富集分析,包括基因本体论(GO)术语和京都基因与基因组百科全书(KEGG)通路。GO功能富集分析包括细胞成分(CC)、生物过程(BP)和分子功能,并使用R软件的ggplot2[3.4.4]包进行可视化分析。
1.1.5 DEARGs的蛋白质‑蛋白质相互作用(PPI)网络构建
利用STRING数据库(https://string‑db.org/)构建DEARGs的PPI网络,使用Cytoscape v3.9.1软件进行可视化分析,最后通过CytoHubba插件筛选10个关键DEARGs。
1.2 动物实验分析
1.2.1 实验动物
选用8~12周龄的无特定病原体级健康C57BL/6雄性小鼠共16只,饲养房间维持室温22 ℃~23 ℃,湿度恒定50%~70%,并应用光照模拟12 h/12 h的昼夜循环,保证适量的饮水和饲料供给。小鼠在建模前采用随机数字表法分为Con组和VILI组,每组8只。
1.2.2 VILI模型构建
两组小鼠实验前均禁食禁饮12 h,模型构建前使用1%戊巴比妥钠麻醉小鼠(腹腔注射40 mg/kg),待小鼠无体动反应后固定小鼠,选用内径为0.5 mm的气管导管行气管插管,呼吸机参数调整如下:潮气量30 ml/kg,呼吸频率100次/min,吸呼比1∶2,呼气末正压0 cm H2O(1 cm H2O=0.098 kPa),吸入氧浓度0.21。VILI组小鼠插管后呼吸机连续通气4 h,Con组小鼠插管后自主呼吸4 h。
1.2.3 苏木精‑伊红(H‑E)染色评估肺组织病理学改变
通气结束,深度麻醉后处死小鼠,随后分离肺组织。将取出的肺组织用多聚甲醛固定,经脱水、浸蜡及包埋后制成切片。切片经脱蜡水化后,依次进行苏木精染色、分化、返蓝及伊红染色,最后脱水透明并用中性树胶封片。染色完成后,利用光学显微镜观察两组小鼠肺组织形态学改变,重点观察肺泡结构的完整性、肺泡壁厚度、是否存在炎性细胞浸润、支气管管壁是否规整、有无间质水肿以及充血等病理特征,以综合评估肺损伤程度。
1.2.4 酶联免疫吸附试验(ELISA)检测肺组织炎症因子白细胞介素(IL)‑1β和肿瘤坏死因子(TNF)‑α的水平
按照IL‑1β和TNF‑α的ELISA试剂盒说明书进行操作,使用酶标仪测定450 nm波长时各孔的光密度值。每组设定4个肺组织样品进行检测,每个样品设定3个复孔以确保数据可靠,最终根据标准曲线计算IL‑1β和TNF‑α的水平。
1.2.5 总RNA提取和逆转录‑实时荧光定量聚合酶链式反应(RT‑qPCR)
使用RNA提取试剂盒提取两组小鼠肺组织的总RNA,并用逆转录试剂盒反转录为互补脱氧核糖核酸,逆转录体系见表1。设计DEARGs、溶酶体标志物溶酶体相关膜蛋白1基因(lamp1)及内参基因β‑肌动蛋白(β‑actin)的特异性引物,由北京擎科生物科技股份有限公司合成,引物序列见表2。使用2×SYBR Green qPCR混合液进行RT‑qPCR。反应条件:95℃预变性30 s、95 ℃变性15 s、65 ℃退火延伸30 s,40个循环。RT‑qPCR反应体系见表3。反应结束后,分析荧光曲线和Ct值。以2−ΔΔCt法计算DEARGs及lamp1的mRNA在两组小鼠肺组织中的相对表达量。



1.2.6 免疫印迹法(WB)检测两组小鼠肺组织中螯合体1(p62/SQSTM1)及微管相关蛋白1轻链3B(LC3B)‑Ⅱ/LC3B‑Ⅰ的水平
为验证筛选出的关键基因sqstm1在蛋白水平的表达及其与自噬过程的关系,本研究采用WB检测了p62/SQSTM1及自噬标志物LC3B的表达。两组小鼠肺组织剪碎后加入蛋白酶抑制剂和裂解液,低温研磨裂解后离心收集上清液。采用二喹啉甲酸蛋白定量试剂盒测定两组小鼠各样品的总蛋白浓度,加入蛋白上样缓冲液,于95 ℃煮沸5 min使蛋白充分变性。将蛋白样品在电泳装置中进行十二烷基硫酸钠‑聚丙烯酰胺凝胶电泳,电泳结束后,转移至聚偏二氟乙烯膜。室温下封闭15 min,随后加入p62/SQSTM1、LC3B和β‑actin一抗于4 ℃孵育过夜,洗膜10 min×3次,加入辣根过氧化物酶标记的羊抗兔二抗,室温下孵育2 h。经含吐温‑20的Tris缓冲盐溶液洗膜10 min×3次后加入化学发光液在暗室条件下显影。通过图像分析软件计算目的蛋白条带灰度值。以目的蛋白p62/SQSTM1与内参蛋白β‑actin的灰度值比值表示p62/SQSTM1的水平;同时,通过计算LC3B‑Ⅱ/LC3B‑Ⅰ以评估两组小鼠肺组织自噬通量的变化。
1.2.7 透射电镜(TEM)观察肺组织中自噬溶酶体改变
用手术刀在培养皿的固定液中将两组小鼠的肺组织切取为1 mm×3 mm的薄片状组织块,厚度尽量控制在1 mm以内,整个过程在1~3 min 内完成,将小组织块离体取下后立即投入预备的装有电镜固定液的培养皿内。将两组小鼠肺组织块置于预冷的2.5%戊二醛溶液中,并在4 ℃下固定24 h,磷酸盐缓冲液漂洗后锇酸固定,脱水、包埋、切片、铀铅双染色后,TEM下观察两组小鼠肺组织中自噬溶酶体变化。
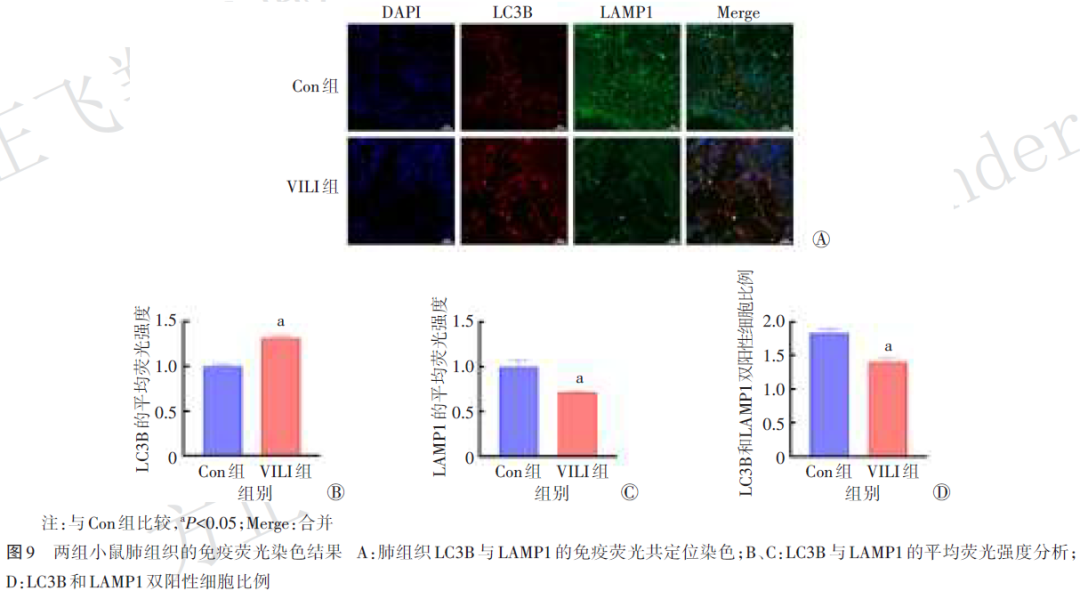
1.2.8 免疫荧光染色检测LC3B与溶酶体相关膜蛋白1(LAMP1)的表达水平及共定位情况
两组小鼠肺组织切片经脱蜡、水化及抗原修复后,使用血清封闭后分别加入抗LC3B和抗LAMP1一抗,于4 ℃孵育过夜。次日洗涤后,加入Alexa Fluor 488标记的羊抗兔二抗和三甲川菁标记的羊抗鼠二抗以分别结合LAMP1与LC3B,避光孵育。细胞核以4',6‑二脒基‑2‑苯基吲哚复染。封片后在激光共聚焦显微镜下观察并采集图像。采用Image J软件测量所取视野内LC3B和LAMP1的平均荧光强度,以定量分析其表达水平;通过LC3B和LAMP1双阳性细胞比例评估共定位情况,以反映两组小鼠肺组织中自噬体与溶酶体的融合效率。以上分析由不知分组的实验人员完成。
2 结 果
2.1 DEGs的筛选
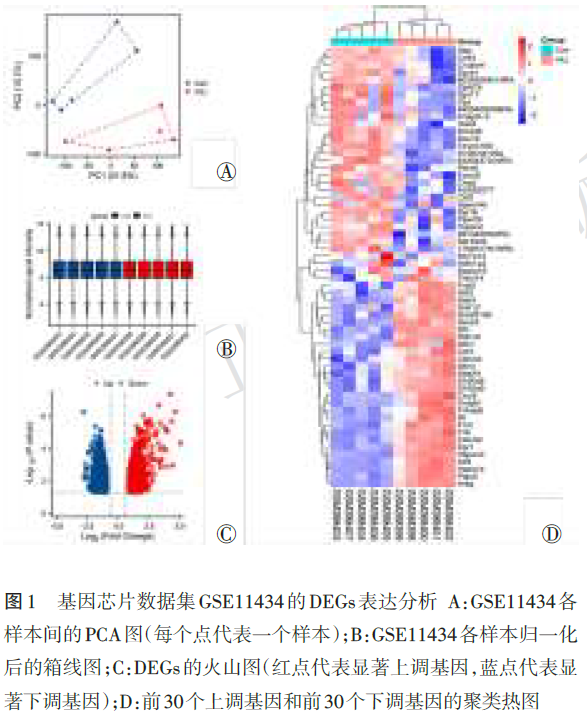
PCA图显示,组内样本聚类良好,组间样本显著分离(图1Ⓐ)。归一化后的箱线图显示各样本中位线分布一致(图1Ⓑ)。火山图显示,共筛选出1 780个DEGs,其中上调的DEGs 944个,下调的DEGs 836个(图1Ⓒ)。聚类热图提示同组内样本的表达高度一致,且组间区分度极大(图1Ⓓ),验证了筛选出的DEGs能够准确代表VILI的病理特征。[<TPL:H:\office\BaiduSyncdisk\杂志\杂志\小样\20265\重拍后飞翔(论著)
2.2 VILI中DEARGs的识别
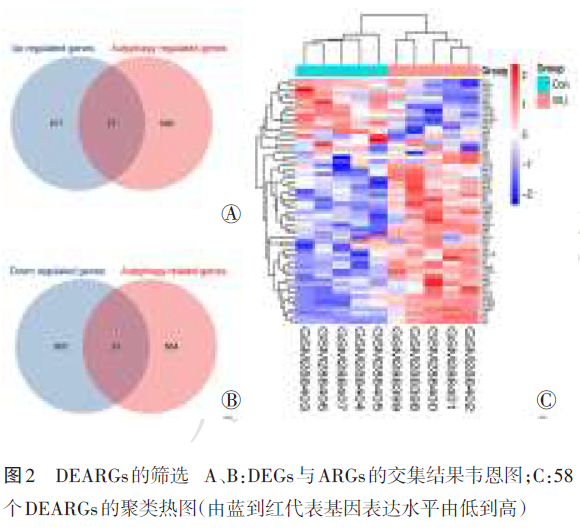
韦恩图结果显示,1 780个DEGs与695个小鼠ARGs进行交集分析后获得58个DEARGs,其中上调基因有27个(图2Ⓐ),下调基因31个(图2Ⓑ)。聚类热图表明,同组内各样本的基因表达趋势基本一致,而两组之间则呈现出不同的空间分布与表达模式(图2Ⓒ)。
2.3 DEARGs的功能富集分析
2.4 PPI网络构建和关键DEARGs的识别
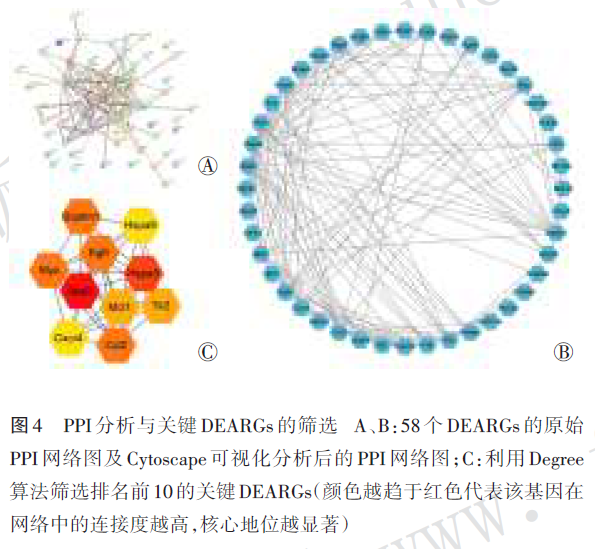
DEARGs的原始PPI网络见图4Ⓐ,对PPI网络的分析见图4Ⓑ,筛选出的10个关键DEARGs见图4Ⓒ。10个关键DEARGs包括stat3、hspa5、ccl2、sqstm1、myc、egfr、tlr2、mcl1、cxcr4和hspa8,均为上调基因。
2.5 两组小鼠肺组织H‑E染色结果及炎症因子IL‑1β、TNF‑α水平
Con组小鼠肺组织H‑E染色切片显示,肺泡结构清晰完整,肺泡壁薄且均匀,无明显渗出物及炎症细胞浸润;各级支气管管壁规整,无明显增厚及破坏;肺间质无水肿、充血表现(图5Ⓐ)。VILI组小鼠的肺组织H‑E染色呈现出肺泡结构紊乱,部分肺泡出现塌陷、融合,肺泡壁明显增厚,可见大量炎症细胞浸润,肺泡腔内有红细胞;支气管管壁也出现不同程度的增厚;肺间质可见明显的充血、水肿(图5Ⓑ)。
ELISA结果提示,与Con组比较,VILI组小鼠肺组织IL‑1β与TNF‑α水平显著升高(均P<0.05,图5Ⓒ、Ⓓ)。
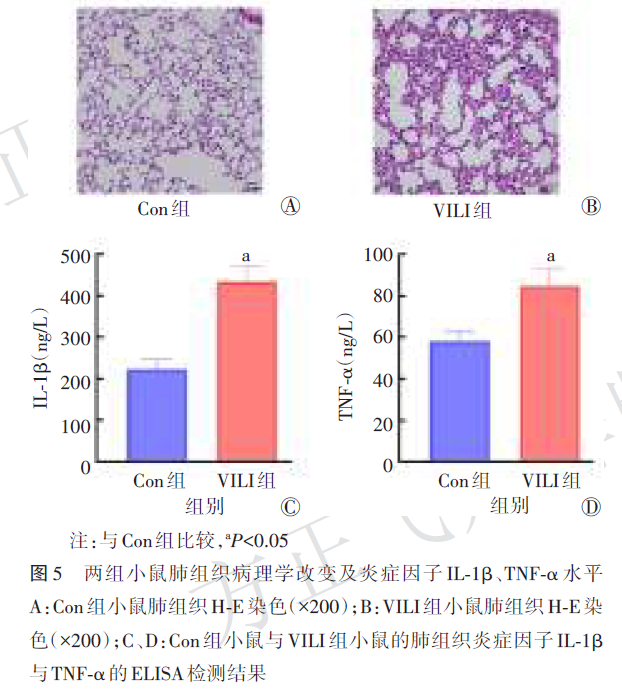
2.6 两组小鼠肺组织DEARGs与lamp1的mRNA相对表达量比较
与Con组比较,10个关键DEARGs中,sqstm1、ccl2、cxcr4、hspa5、hspa8、myc、stat3、tlr2的mRNA相对表达量在VILI组小鼠肺组织中均明显升高(均P<0.05,图6)。与Con组比较,lamp1的mRNA相对表达量在VILI组小鼠肺组织中显著降低(P<0.05,图7)。
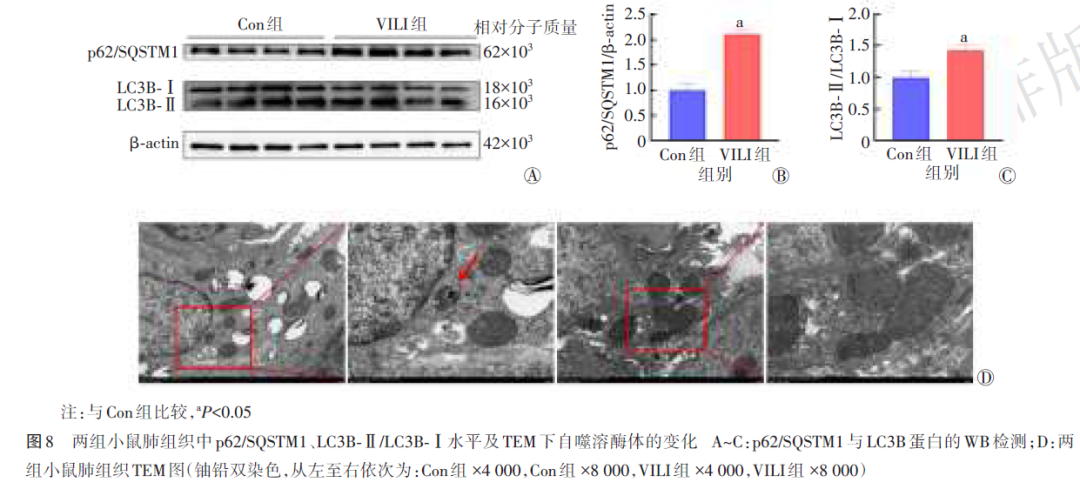
2.7 两组小鼠WB及TEM检测结果比较
WB结果显示,与Con组比较,VILI组小鼠肺组织p62/SQSTM1水平显著升高(P<0.05),同时LC3B‑Ⅱ/LC3B‑Ⅰ亦升高(P<0.05)(图8Ⓐ~Ⓒ)。TEM结果显示,Con组小鼠肺组织中可见数量及形态正常的自噬溶酶体,而VILI组小鼠肺组织中几乎未见自噬溶酶体(图8Ⓓ)。
2.8 两组小鼠肺组织LC3B和LAMP1表达水平比较与共定位分析
与Con组比较,VILI组小鼠肺组织中LC3B荧光强度明显增强(P<0.05),同时LAMP1荧光强度明显减弱(P<0.05,图9Ⓐ~Ⓒ)。LC3B与LAMP1的共定位结果表明,与Con组比较,VILI组小鼠肺组织中LC3B和LAMP1双阳性细胞比例显著减少(P<0.05,图9Ⓓ)。
3 讨 论
本研究利用生物信息学分析筛选DEARGs并构建动物模型,在VILI小鼠模型中观察到肺组织发生显著的病理损伤,伴随炎症因子IL‑1β、TNF‑α水平的显著升高;RT‑qPCR结果显示,与Con组小鼠比较,sqstm1、ccl2、cxcr4、hspa5、hspa8、myc、stat3及tlr2的mRNA相对表达量在VILI组小鼠肺组织中显著升高;为进一步验证筛选出的关键基因sqstm1在蛋白水平的表达及其与自噬过程的关系,本研究采用WB检测了p62/SQSTM1及自噬标志物LC3B的表达,结果显示,与Con组小鼠比较,VILI组小鼠肺组织p62/SQSTM1水平显著增加,且LC3B‑Ⅱ/LC3B‑Ⅰ呈现升高趋势;TEM结果显示,Con组小鼠肺组织中可见数量及形态正常的自噬溶酶体,而VILI组小鼠肺组织中几乎未见自噬溶酶体;为明确自噬体与溶酶体融合异常在VILI自噬通量障碍中的作用,本研究利用RT‑qPCR检测了两组小鼠肺组织中溶酶体标志物lamp1的mRNA相对表达量,结果提示,与Con组比较,VILI组小鼠肺组织中溶酶体标志物lamp1的mRNA相对表达量显著降低;免疫荧光染色结果表明VILI组小鼠肺组织中LC3B平均荧光强度显著增强,LAMP1平均荧光强度显著减弱,且LC3B和LAMP1双阳性细胞比例显著减少,提示VILI组小鼠肺组织中自噬体与溶酶体融合异常,符合自噬通量障碍的特征。这些结果提示自噬通量障碍可能为VILI的重要病理环节。
本研究关注到,在自噬状态的动态评估中,单一指标的升高往往具有多义性。本研究观察到LC3B‑Ⅱ/LC3B‑Ⅰ与p62/SQSTM1水平呈现同步升高的特征。本研究利用TEM在超微结构水平进一步验证了这一推测,结果显示,VILI组小鼠肺组织中几乎未见自噬溶酶体。考虑到自噬体作为中间产物,其半衰期极短(仅10~20 min),在静态断面中捕捉到典型双膜自噬体的概率相对较小;相比之下,作为降解场所的自噬溶酶体若发生缺失,则是自噬体与溶酶体融合障碍的强有力间接证据。本研究观察到的自噬溶酶体显著减少与p62/SQSTM1水平升高互为验证,共同揭示了自噬通量障碍可能是VILI发生过程中的核心病理特征。RT‑qPCR结果提示VILI组小鼠肺组织lamp1的mRNA相对表达量较Con组显著降低;免疫荧光染色结果显示,与Con组比较,VILI组小鼠肺组织中LC3B平均荧光强度显著增强,LAMP1平均荧光强度显著减弱,并伴随LC3B和LAMP1双阳性细胞比例显著减少。这些结果提示VILI小鼠肺组织不仅存在自噬体的病理性积累,还伴随溶酶体功能受损,从而导致自噬体与溶酶体的融合过程障碍。这从细胞生物学角度进一步解释了前述p62/SQSTM1蓄积的原因:即由于融合环节的障碍,导致自噬体无法转化为自噬溶酶体进行最终降解。
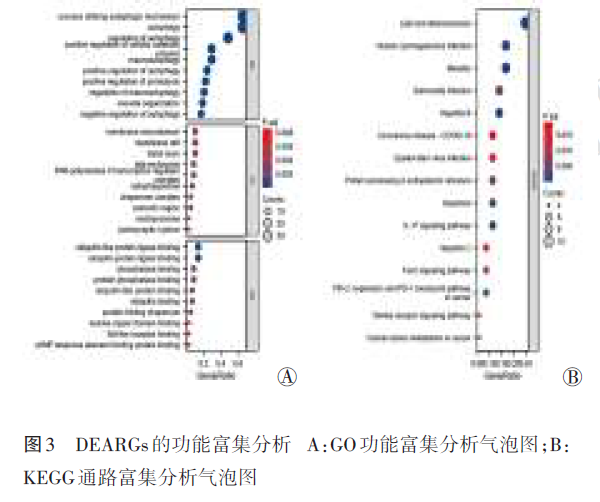
对于DEARGs的功能富集分析,从GO功能富集分析结果看,BP主要聚焦于“自噬、利用自噬机制的过程、自噬的调节等”,CC主要定位于“自噬体、胞质区、分子伴侣复合体等”,揭示了DEARGs在介导自噬体形成、溶酶体降解及蛋白质稳态维持中的关键作用。KEGG通路富集分析结果虽以感染和免疫相关通路为主,但这些通路正是自噬‑炎症轴的重要上游,“麻疹、乙肝与巨细胞病毒”均可诱导并重塑自噬过程,其可通过与曲螺旋肌球蛋白样BCL2相互作用蛋白1特异性结合,从而阻断自噬末端融合阶段来增加复制。IL‑17信号通路作为中性粒细胞主导炎症的关键驱动,也可使自噬过程失调,从而形成“细胞因子‑自噬‑炎症”的调控机制;功能富集条目“脂质和动脉粥样硬化”则提示了巨噬细胞自噬溶酶体的形成与炎症小体调控的关联,提示激活该轴可抑制炎症。此外,“脂质和动脉粥样硬化”还提示了自噬‑溶酶体途径对核苷酸结合寡聚化结构域样受体蛋白3炎症小体的负向调控作用;结合本研究的发现,激活该调控轴可能成为抑制VILI炎症级联反应、减轻组织损伤的潜在策略。
本研究筛选并鉴定出8个与VILI发病及自噬调控密切相关的DEARGs。针对这些分子或通路在自噬过程中发挥的具体作用,后续可开展体外模型或在体实验进一步验证。
本研究中egfr与mcl1虽为DEARGs,但经RT‑qPCR验证未发现显著的差异表达,这可能是因为egfr和mcl1的表达变化发生在复氧后更晚的时间点,而本研究取材的时间不在其变化的峰值区间;也可能因其只在某些特定细胞类型中差异表达,所以在整体肺组织的分析中未能见到变化。
本研究后续还可通过转染单体红色荧光蛋白‑增强型绿色荧光蛋白‑LC3腺病毒的方法,并结合溶酶体抑制剂或融合阻断实验,以进一步精确分析自噬体与溶酶体融合障碍的分子细节。此外,本研究所用的通气参数(潮气量≈30 ml/kg,4 h)与数据集GSE11434中所用的通气参数(潮气量≈20 ml/kg,3 h)存在一定差异。通气参数的差异可能对自噬通量及基因表达产生一定影响,后续研究如果能统一通气参数,将更有利于机制验证。
国际麻醉学与复苏杂志,2026,47(05):398-409 .
DOI:10.3760/cma.j.cn321761-20250924‑00452
小提示:本篇资讯仅在梅斯医学APP中开放阅读,请扫描二维码直接下载APP
